
 | |
Odkąd odkryto silną zależność pomiędzy snem REM a marzeniami sennymi, wzrosło zaciekawienie naukowców na temat mechanizmów odpowiedzialnych za procesy śnienia. Na podstawie badań neurofizjologicznych uważa się, że sygnały powstające w wyniku aktywacji pewnych neuronów części mózgu zwanej mostem, dochodząc do ośrodków sensorycznych w korze mobilizują ją do syntezy wrażeń zmysłowych, odbieranych jako mniej lub bardziej spójne struktury narracyjne, czyli nasze sny. Jednocześnie wyjście motoryczne jest zablokowane dzięki atonii mięśni w wyniku działania mechanizmów zlokalizowanych na poziomie pnia mózgu, co zabezpiecza nas przed odgrywaniem śnionych ruchów fizycznym ciałem (chyba, że cierpimy na RBD). Sen REM jest także związany ze wzrostem przepływu krwi w obszarze wzrokowych pól kojarzeniowych oraz obszarów limbicznych oraz ze zmniejszeniem metabolizmu w obszarach grzbietowo-bocznej kory przedczołowej (DLPFC), pierwszorzędowej kory wzrokowej i przedklinka. Uważa się, że taki a nie inny wzorzec aktywacji odpowiada za pewne cechy marzeń sennych jak wyraźne wrażenia wizualne, doświadczanie silnych emocji oraz brak refleksji, krytycznej oceny i wolnej woli.
Jednak bardziej szczegółowe badanie neuronalnych korelatów konkretnych elementów marzeń sennych wciąż stanowi problem. Tradycyjne metody obrazowania snów nie potrafią bowiem odnieść się do określonych wydarzeń ze snu, gdyż jak dotąd z treścią marzenia sennego można było się zapoznać dopiero po obudzeniu badanego, a więc po fakcie. Co więcej, sprawozdanie ze snu stanowiło subiektywny opis wydarzeń, pełen niewyraźnych lub niepełnych śladów pamięciowych, niemożliwych do dokładnego umiejscowienia ich w czasie. Dopiero zastosowanie narzędzia umożliwiającego określenie precyzyjnych ram czasowych dla wykonywanych we śnie czynności dałoby możliwość dokładnego odniesienia danych zebranych podczas snu za pomocą technik obrazowania do poszczególnych elementów marzenia sennego. Aby to zrobić najlepiej byłoby informować w czasie rzeczywistym o tym, co w danej chwili robimy we śnie, cały czas w nim pozostając.
Właśnie do tego możemy zastosować stan świadomego snu, co już wielokrotnie udowadniano. Jednak tym razem, po raz pierwszy, zastosowano świadome śnienie w połączeniu z podstawowymi i jednocześnie najnowocześniejszymi technikami obrazowania śpiącego mózgu (polisomnografia, fMRI, NIRS). W badaniu z „Current Biology” skupiono się na neuronalnych korelatach konkretnego elementu marzenia sennego, jakim było zaciskanie pięści, na zmianę, lewej i prawej ręki. Jak w wielu innych badaniach z wykorzystaniem świadomych snów, także i w tym użyto predefiniowanych sekwencji ruchu gałek ocznych, lewo-prawo-lewo-prawo (LPLP), które miały posłużyć za czasowe markery oznaczające początek świadomego snu, oraz rozpoczęcie i zakończenie każdej doświadczalnej serii wykonywanej czynności. Jak to działa? Obok mięśni oddechowych, mięśnie poruszające oczami nie podlegają pod mechanizmy odpowiadające za paraliż podczas snu REM, co więcej wykazano także, iż kierunek ruchów gałek ocznych podczas snu odzwierciedla kierunek, w którym śniąca osoba spogląda, obserwując mające w tym czasie miejsce we śnie wydarzenia (tzw. scanning hypothesis). W każdym badaniu związanym ze snem stosuje się tzw. elektrookulogram (EOG), który zapisuje aktywność mięśni okołoocznych, a więc i zapis ruchu oczu – ponadto, z takiego zapisu możemy odczytać, czy oczy poruszały się w lewo czy w prawo. Zatem jeżeli podczas snu będziemy wiedzieli, że śnimy, i będziemy pamiętać o tym, że mieliśmy wykonać jakieś zadanie poprzedzone ustalonym sygnałem, będziemy mogli zaznaczyć dowolny moment snu za pomocą wcześniej ustalonych sygnałów, którymi będą właśnie określone ruchy gałek ocznych.
Badani (osoby z dużym doświadczeniem w świadomym śnieniu) zostali poinstruowani, aby po osiągnięciu stanu świadomego snu zasygnalizowali ten moment sygnałem LPLP, po czym natychmiast wykonali zaciskanie pięści lewej dłoni. Następnie mieli ponownie zasygnalizować LPLP i wykonać to samo, z tym że drugą ręką. Czynności te były powtarzanie tak długo jak było to możliwe, przy czym za każdym razem sygnalizowano zmianę rąk ustalonym sygnałem ocznym. W tym samym czasie badanym mierzono sygnał EMG (aktywność elektryczna mięśni) i EEG, który wskazywał na przebywanie we śnie REM, podczas gdy EOG rejestrowało wolicjonalne sygnały z mięśni ocznych, które były wyraźne odróżnialne na tle pozostałych sygnałów charakterystycznych dla tej fazy snu (dla przypomnienia, nazwa snu REM pochodzi od ang. rapid eye movement, a więc od szybkich ruchów gałek ocznych, które są cechą charakterystyczną tego stadium). Jednocześnie przeprowadzono rejestrację aktywności mózgu za pomocą funkcjonalnego rezonansu magnetycznego (fMRI) lub spektroskopu w bliskiej podczerwieni (NIRS).
Chociaż w badaniu brało udział 6 osób, planowane zadanie wykonały jedynie dwie osoby: jedna z nich badana była za pomocą fMRI, a druga – NIRS. Jako pomiar kontrolny, te same osoby wykonywały zaciskanie pięści w dwóch kolejnych wariantach, oba w stanie czuwania: wykonując fizyczne ruchy oraz wyobrażając sobie zaciskanie pięści.
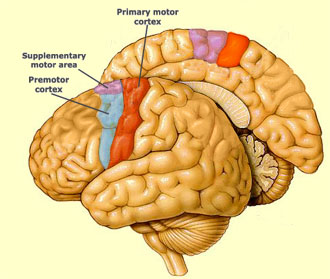 | |
| Premotor cortex = kora przedruchowa Primary motor cortex = pierwszorzędowa kora ruchowa Supplemetary motor area = dodatkowe pole ruchowe |
Wynik tych badań pozwalają także rzucić nieco światła na sprawę kontrowersji dotyczącej miejsca kontroli procesów hamowania wyjścia motorycznego podczas ruchów wykonywanych w wyobraźni, jak i we śnie. O ile wiadomo, że atonia podczas snu generowana jest na poziomie pnia mózgu i rdzenia kręgowego, nie do końca wiadomo jest czy hamowanie egzekucji ruchów wyobrażanych odbywa się na poziomie korowo-korowym, tzn. zanim informacja z kory przedruchowej przejdzie do pierwszorzędowej kory ruchowej, wyjście motoryczne prowadzące do wykonania fizycznego ruchu jest hamowane, przy czym źródłem tej inhibicji jest kora przedczołowa. Jednak podobieństwo, jakie zaobserwowano także pomiędzy wzorcem aktywacji podczas wykonywania ruchów wykonywanych świadomie podczas snu i tych wykonywanych w wyobraźni, potwierdza w jakiś sposób hipotezę o tym, że instrukcje dotyczące inhibicji motorycznej nie pochodzą z kory przedczołowej, lecz sygnał z kory ruchowej jest przekazywany do motoneuronów rdzenia, gdzie dopiero wtedy następuje hamowanie czynności ruchowych. Gdybyśmy mieli do czynienia z hamowaniem korowo-korowym, nie powinno być aktywacji pierwszorzędowej kory ruchowej, ani podczas ruchu wyobrażanego, ani śnionego.
Autorzy badań podkreślają, że jest to badanie typu proof-of-concept, a więc mające na celu udowodnienie poprawności pewnej idei, w tym wypadku – metody badawczej, umożliwiającej odczytywanie treści marzeń sennych, na podstawie aktywności mózgu. Jest to wprawdzie początkowa, ale niezwykle istotna faza badań nad snami prowadząca do opracowania nowych rozwiązań „czytania” snów.
Miejmy również nadzieję, że badanie to zachęci innych naukowców, zajmujących się snami i świadomością, aby w swoich eksperymentach zaczęli wykorzystywać świadome śnienie, jako dodatkowe narzędzie badawcze.
Dresler M, Koch SP, Wehrle R, Spoormaker VI, Holsboer F, Steiger A, Sämann PG, Obrig H, & Czisch M (2011). Dreamed Movement Elicits Activation in the Sensorimotor Cortex. Current biology : CB PMID: 22036177
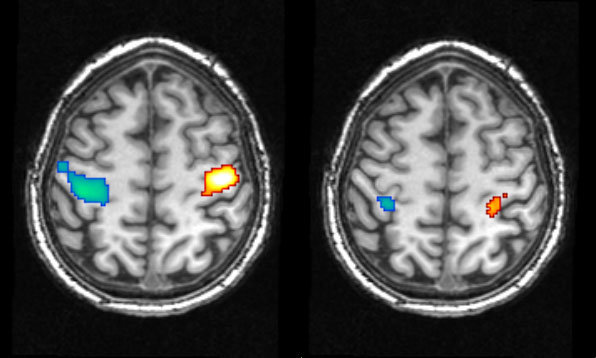
Foto: Dresler i in. 2011, aktywność w obszarze kory ruchowej w trakcie zaciskania pięści na jawie (po lewej) oraz podczas świadomego snu (po prawej). Niebieski kolor oznacza aktywność podczas ruchów wykonywanych prawą ręką (dlatego widzimy ją w lewej półkuli), a czerwony - lewą ręką (aktywność w prawej półkuli mózgu). Objaśnienia w tekście.







Mam uwagę do pierwszego zdjecia z obrazami z fMRI - jeśli jest to standardowo preprowadzane badanie obrazowe, to to co widzimy na obrazie po lewej, to prawa strona badanego (leży on jakby "nogami do nas"). W takim układzie niebieski kolor dotyczy prawej półkuli/lewej ręki.
OdpowiedzUsuńna tym zdjeciu lewa strona mozgu jest lewą półkula w rzeczywistości. góra zdjecia- płaty czołowe, dół- czesc potyliczna. widac zresztą po kształcie mózgu.
OdpowiedzUsuńMuszę przyznać, że są to naprawdę bardzo ciekawe spostrzeżenia. Nie miałam pojęcia, że nasze sny mają taką moc i nasz mózg tak na nie reaguje. Szkoda, że nie wszystkie pamiętamy, nie mniej pojawiają się takie również o których ciężko zapomnieć.
OdpowiedzUsuńSkoro już mowa o snach, to myślę, że bardzo ważnym problemem jest narkolepsja, czyli nadmierna senność. Bardzo uciążliwa i przede wszystkim męcząca choroba, dlatego szuka się różnych sposobów na zwalczenie jej. Odsyłam Was na stronę https://baza-lekow.com.pl/nadmierna-sennosc-czyli-czym-jest-narkolepsja/ , na której znajdziecie sobie szczegółowe informacje dotyczące tych zaburzeń.
OdpowiedzUsuńDobrze wiedzieć o takich kwestiach.
OdpowiedzUsuńFakt, problemy ze snem to poważny problem z jakim boryka się dużo osób. Można to w zasadzie uznać za chorobę cywilizacyjną. Przyczyn tego stanu rzeczy może być wiele, zaczynając od stresującej pracy, a kończąc na złej diecie. Dlatego kluczowa w leczeniu będzie diagnoza specjalisty oraz skuteczne leczenie. Często polecane są naturalne tabletki na sen - https://www.boiron.pl/nasze-produkty/leki-zlozone/sedatif-pc przy stanach nadmiernej pobudliwości nerwowej.
OdpowiedzUsuńDlatego właśnie badania rezonansu magnetycznego są niesamowite. W bardzo prosty i mało inwazyjny sposób możemy zajrzeć do wnętrza ciała i obserwować zmiany. Dzięki temu leczenie chorób lub urazów jest o wiele skuteczniejsze, a niektóre choroby wcześnie wykryte można szybko i skutecznie wyleczyć.
OdpowiedzUsuń